Axonogenesis
Axon is truly unique to neurons. Axon is the morphological hallmark of neurons. Its formation establishes neuronal polarity and is prerequisite to synapse formation and circuit wiring, both of which underlie cognition and behaviours. Axon degeneration is one indicator for various neurodegenerative diseases and brain injury. Therefore, tremendous interest and effort have been placed on understanding and inducing axon regeneration from learning developmental axonogenesis.
Even with substantial description of cell biology and molecular players of axon formation, we still know little about genetic control of axonogenesis. For example, less work has been done to understand why only neurons and no other cell types have an axon. Studies of axon formation have substantially borrowed and benefited from findings of planar cell polarity, emphasizing the similarity of their mechanisms. However, axonogenesis encompasses more complicated aspects than establishing cell polarity, and planar cells never grow an axon. To ensure axon production, one possibility is that neurons are uniquely equipped with a set of special regulatory rules.
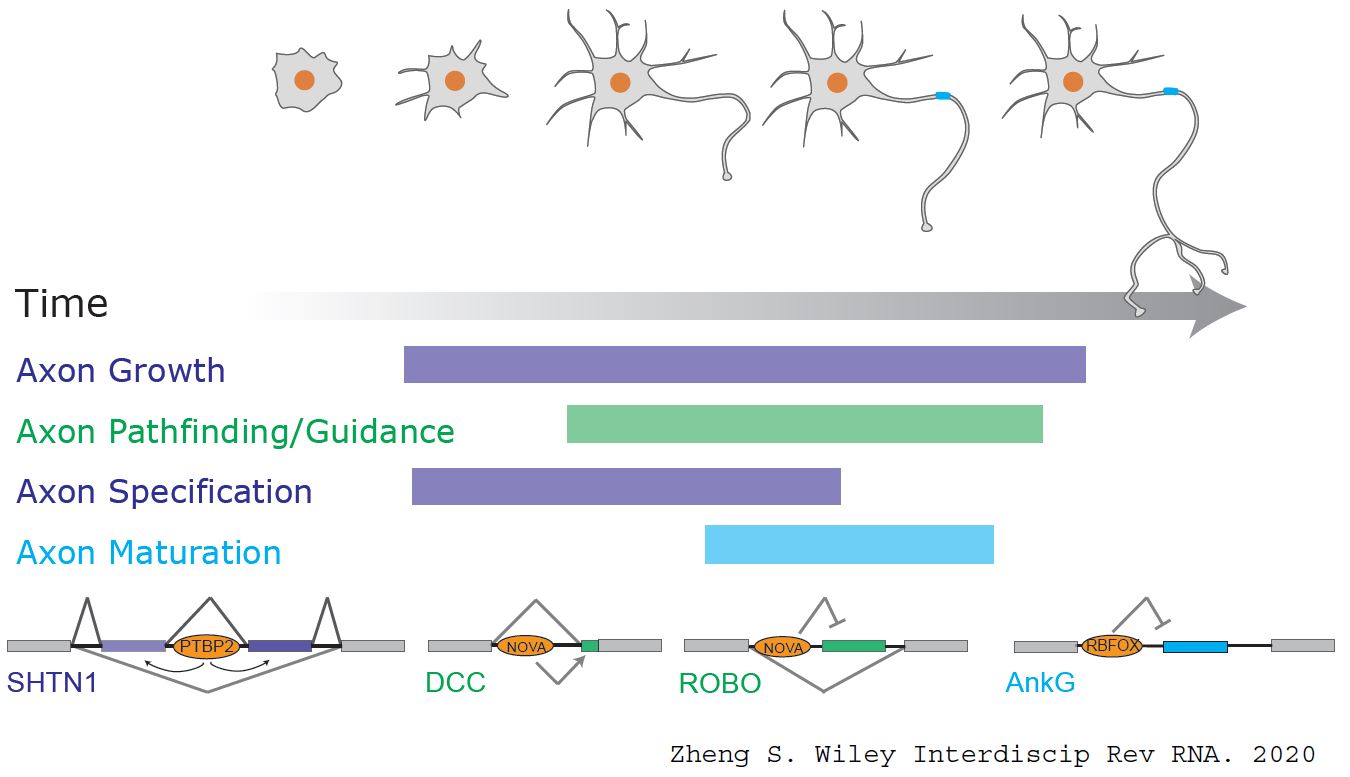
We found early axonogenesis is coordinated by neural-specific splicing through unbiased transcriptome analysis (Zhang et al., 2019). Many genes change isoform expression during this period and these alternative splicing events exhibit neural-specific splicing patterns, raising the possibility that neuron-specific splicing may be part of the cell type specific regulatory rules enabling axon formation exclusively in neurons. Furthermore, known axonogenesis genes do not change gene expression when a neuron grows out an axon, but many change splicing (Zhang et al., 2019). Therefore, their contributions to axonongenesis are coordinated at the splicing level. A reasonable speculation is that these genes adopt new functions through alternative splicing that occurs only in neurons to take on the task of neuron-exclusive axon formation.
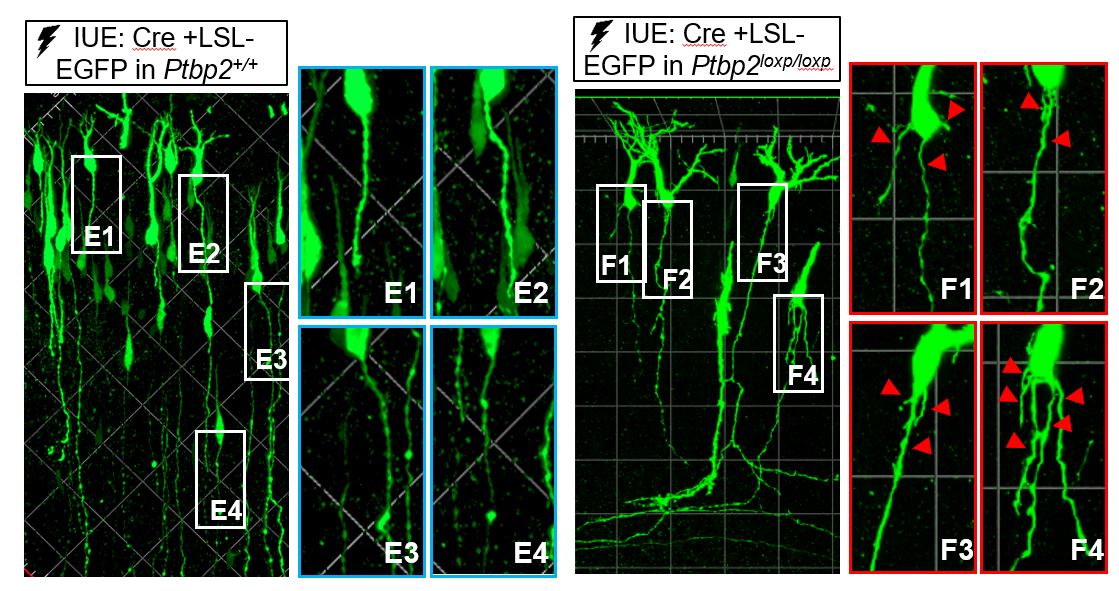
We identified PTBP2 as a regulator of axonogenesis-associated splicing. PTBP2 depletion impedes axonal growth and simultaneously stimulating axon specification in vitro and in vivo. While Ptbp2-/- axons are generally shorter than wildtype axons, some Ptbp2 null neurons can extend two axons with molecular markers of developed axons: TAU1 and ANKG. These two distinct phenotypes demonstrate the inequality between axon growth and specification. One possible explanation for the Ptbp2-/- phenotype is decreased expression of isoforms for axon growth and increased expression of isoforms for axon specification. In support of this hypothesis, Shtn1 splicing is regulated by PTBP2 such that SHNT1L is decreased and SHTN1S is increased in the knockout neurons. Compensation for the SHNT1L shortage partially rescues the axon length in Ptbp2-/- neurons. Therefore, alternative splicing regulation during early axon formation is partly to ensure balanced expression of the isoforms responsible for two integrated aspects of early axonongenesis.